другая биологическая (растительная) структура, веточка кустарника, находящаяся в процессе увядания. Отсеченная от корневой системы веточка кустарника (передавшая система) из-за недостатка влаги в соответствии с собственной функцией диссипации переходит в наиболее вероятное состояние: гибели или увядания. И, соответственно, в окружающее пространство Lвыбрасывает¦ информационный поток определенной интенсивности. (Н.А. Козырев это связывал с понятием излучения времени.) Точка 2 на рисунке N 1 является началом информационного влияния. Информационное воздействие продолжалось в течении 3000 с, на рисунке N 1 ему соответствует участок 2-6.
Увядающая веточка кустарника использованная в качестве источника информационного потока, в течение всего времени влияния (участок 2-6, рисунок N 1) находилась в пустой (сухой) емкости для воды*. Смысл эксперимента заключался еще и в том, чтобы в произвольно выбранный момент времени добавить в пустую емкость столь, необходимую для увядающей структуры влагу, прекратив тем самым процесс увядания. Более того, заменить его обратным процессом, структурированием той же самой биологической структуры. Таким образом, биологический процесс, идущий с увеличением собственной энтропии биоструктуры и излучающий информацию, сменится на обратный, характеризующийся сокращением этого определяющего параметра, и как прямое следствие этого - изменится и направление информационного влияния (обмена). Что особенно важно - источником информационных влияний различных знаков оказывается одна и та же биологическая структура и это показательно. Таким моментом времени является точка 6 на рисунке N 1. а участок 6-21 показывает реакцию рецептора на происходящий в непосредственной близости от него биологический процесс структурирования источника информационного влияния (веточки кустарника).
Точка 21 представляет в свою очередь момент времени, когда вся Lсистема информационного влияния¦ была вынесена из пространственной области эксперимента, то есть процесс информационного влияния был прекращен, участок 21-30 (рисунок N 1) - послеэкспериментальный фоновый дрейф параметра Gh.
Попытаемся проанализировать полученные результаты. Участок 1-2 соответствует фоновому дрейфу контролируемого параметра Gh, и может быть истолкован как реакция использованной рецепторной системы, некоторого объема растительной ткани, на происходящие в природе информационные взаимодействия. После внесения в пространственную область эксперимента источника информационного влияния (точка 2), веточки кустарника, график демонстрирует существенное снижение контролируемого параметра Gh. Вследствие чего рецепторная система обнаруживает снижение собственной энтропии биоструктуры, а это свидетельствует о притоке к ней некоторого количества информации, Этот момент является очень важным, так как характеризует сам механизм информационного обмена в природе между передающими системами, увеличивающими собственную энтропию в силу определенных обстоятельств, и рецепторов, способных воспринимать высвобождающуюся в пространстве информацию. Однако информационный обмен подразумевает и иной знак взаимодействия Lпередающей¦ и рецепторной систем, когда они как бы меняются местами, а рецепторная система сама вынуждена расставаться с некоторым количеством информации под влиянием процессов, идущих с сокращением энтропии. Такой характер информационного обмена демонстрирует участок 6-21. Как уже отмечалось, в ходе эксперимента в некоторый момент времени (точка 6) передающей системе была предоставлена возможность уменьшить свою собственную энтропию структуры. После точки 6 произошла смена знака течения биопроцесса (функционирования) биосистемы информационного влияния увядания на структурирование. После чего график на участке 8-21 демонстрирует изменение реакции на данное событие рецепторной системы. В начале участка имело место некоторое увеличение контролируемого параметра Gh рецепторной системы. Это говорит о том, что информационный поток от передающей системы прекратился.
Более того, контролируемая нами рецепторная система сама вынуждена расставаться с некоторым количеством информации под влиянием процесса структурирования веточки кустарника. Системы, говоря буквально, сменили роли! В дальнейшем наблюдается некоторое сокращение параметра Gh. Воздержимся от комментариев этого явления, но в дальнейшем происходит плавное увеличение контролируемого параметра, свидетельствующего об увеличении рецепторной системой собственной энтропии структуры. Данный процесс имеет место вплоть до момента времени 21, когда вся передающая биосистема (веточка кустарника) была вынесена из пространственной области эксперимента. Дальнейшее поведение графика на участке 21-30 соответствует участку 1-2 с той лишь разницей, что измерения на этом участке проводились после эксперимента. Обращает на себя внимание реакция рецепторной биосистемы на факт удаления из пространственной области эксперимента биосистемы информационного влияния, имевшая место в заключительной фазе эксперимента. Можно проследить на рисунке N 1 некоторое увеличение характеризующего параметра Gh непосредственно после удаления биопроцесса структурирования веточки кустарника. Это указывает на увеличение энтропии рецепторной биоструктуры. Чем вызвана такая ее реакция? Возможным ответом служит предположение о том, что явление передачи некоторого количества информации рецепторной системой под информационным влиянием структурирующейся биосистемы (веточки кустарника) в какой-то степени является добровольным актом рецепторной биосистемы. Поэтому удаление из пространственной области эксперимента биосистемы, на данном его этапе поглощающей некоторое количество информации, расценивается рецепторной биосистемой (во всяком случае - в первые мгновения после удаления биопроцесса структурирования) как внезапное, вызванное какой-то внешней причиной, сокращение ее (веточки кустарника) жизненных сил (биоактивности). И рецепторная система предпринимает усилия, направленные на помощь уже несуществующему собрату.
Здесь необходимо подвести некоторые итоги. Прежде всего, вышеописанный эксперимент показал принципиальную возможность использования некоторого объема растительной ткани в качестве рецептора неэлектромагнитного информационного сообщения (как впрочем, и само его существование). Удалось выявить отличные реакции применявшегося рецептора на информационные влияния различных знаков и обнаружить имеющиеся закономерности. Можно предположить , что растительная биосистема под влиянием определенных причин, непрерывно увеличивающая собственную энтропию, излучает в пространство некоторую информацию. В свою очередь, находящиеся в непосредственной близости от источника информационного сообщения растительные биосистемы способны ее поглощать из пространства, обнаруживая это через сокращение собственной энтропии.
В то же время растительные биосистемы, функционирующие по закону сокращения собственной энтропии, потребляют информацию из пространства, при этом у находящихся рядом других растительных биосистем наблюдается потеря организованности структур.
Однако выявленные в ходе эксперимента характеристики информационного обмена являются недостаточными для понимания глобального информационного взаимодействия в природе. Так, мы использовали в качестве рецепторной системы растительную биосистему, находящуюся в состоянии равновесия, то есть не излучающую и не поглощающую информацию из окружающего ее пространства, если так можно сказать, по собственной инициативе. И каким образом будет протекать информационный обмен, например, двух биосистем, характер функционирования которых в силу определенных причин взаимоотношения со средой произрастания обусловлен увеличением их собственных энтропии и таким образом излучающих информацию в пространство? Как. впрочем, и одновременно поглощающих информацию из пространства (фона)?
Для того, чтобы разобраться в данном вопросе, следует выяснить, от чего зависит количество излучаемой или поглощаемой информации некоторой растительной биоструктурой.Попытаемся сформулировать основные признаки интенсивности возможного информационного влияния со стороны растительной биоструктуры. Предположим, что данное понятие определяется, во-первых: массой (объемом) растительной биоструктуры, участвующей в информационном влиянии, то есть функционирующей по определенному закону изменения собственной энтропии. Во-вторых: вероятно, в равных объемах (массах) различных биоструктур (разных видов растительности) содержится неодинаковое количество Lхранящейся¦ в них информации. (Мы не будем здесь рассматривать характеристики информации, свойственной отдельно взятому виду биоструктуры). И, наконец, в-третьих: необходимо учитывать, что различные растительные биоструктуры (виды растительности) имеют различные функции диссипации, то есть различные скорости перехода как в наиболее вероятное состояние под влиянием поражающих факторов, так и структурирования вследствие благоприятных условий произрастания, и это неминуемо повлияет в целом на количество излучаемой или поглощаемой информации биоструктурой за единицу времени. В то же время следует иметь в виду и то - что второй и третий признаки интенсивности..., возможно каким-то образом, связаны между собой.
Опираясь на вышеизложенное, можно выработать условия проведения эксперимента, который бы решил поставленную ранее задачу.
Однако вначале оценим с позиций сформулированных нами признаков ранее описанный эксперимент, в котором рецепторная система находилась в состояния равновесия и не обнаруживала изменений энтропии собственной биоструктуры. В тоже время значительно превосходящая ее по объему (массе) биосистема информационного влияния формировала информационный поток соответствующей интенсивности.
Имеет смысл провести эксперимент, в котором рецепторная система (PC) является значительно превосходящей по объему (массе) биосистему влияния, и, кроме того, также, как и биосистема влияния, функционирующая по закону увядания (к состоянию гибели). Основной задачей определяем обнаружение характера (направления) информационного обмена между данными биосистемами. Необходимо отметить и тот факт, что несмотря на требуемое условиями эксперимента значительное превосходство в количестве вещества (массе) рецепторной биосистемы над биосистемой информационного влияния (возмущения) межэлектродное пространство (объем растительной ткани между зажимами-электродами) зондового измерителя биоактивности в настоящем эксперименте соответствует аналогичному значению вышео-писанного эксперимента. Кроме того. в ходе предыдущего эксперимента рецепторная биосистема помещалась в свето- и влаго- непроницаемый полиэтиленовый пакет в целях достижения стабилизации контролируемого параметра низкочастотной электропроводности (Gh, наступающей по истечению некоторого времени после помещения рецептора в данный замкнутый объем. В данном эксперименте биосистема рецептора также помещалась в замкнутый объем с той лишь разницей, что проведение процесса информационного обмена был приурочен к моменту до стабилизации контролируемого параметра Gh. Условием эксперимента являлось также и использование в качестве рецепторной и биосистемы влияния одного и того же вида растительности.
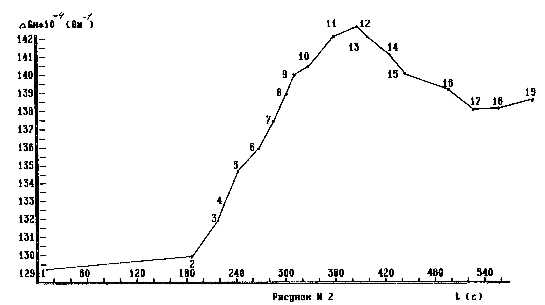
Увеличение параметра Gh рецепторной биосистемы характеризует постоянный рост ее собственной энтропии в ходе продолжающегося (после разделения с корневой системой) процесса увядания. Таким образом¦ участок 1-2 на рисунке N 2 (таблица N 2) представляет фоновый процесс, характеризующийся увеличением энтропии рецепторной биосистемы и имевший место до информационного обмена. Точка 2 (рисунок N 2) является моментом времени установления информационного влияния (внесение биосистемы влияния в пространственную область эксперимента). Участок 2-11 (рисунок N 2) демонстрирует реакции рецепторной биосистемы на заданное информационное влияние. Информационный обмен был прекращен в момент времени 11 (рисунок N 2) - вынос биосистемы влияния из пространственной области эксперимента. Анализ полученных результатов, демонстрируемых рисунком N 2, показывает, что после установления информационного влияния на рецепторную биосистему она обнаружила некоторое увеличение скорости изменения характеризую-щего параметра Gh по сравнению с участком 1-2 предшествующего информационному обмену. Следовательно, биосистема информационного Lвлияния¦ в ходе эксперимента поглощала некоторое количество информации из рецептора. В чем причина подобного информационного взаимодействия? Вероятно, основополагающим фактором, определяющим направление информационного обмена между данными биосистемами, является интенсивность выбрасываемой каждой из биосистем в отдельности в пространство информационного потока.
Учитывая тот факт, что рецепторная биосистема имела преимущества в количестве вещества, участвующего в информационном обмене, и принимая во внимание, что обе биосистемы представлены одним видом растительности, можно предположить,
Таблица № 2
Номер измерения (рис, N 2) |
DG*10-4(om-1) |
Время (с) |
1 |
128,5 |
0 |
2 |
130 |
190 |
3 |
132 |
220 |
4 |
133 |
230 |
5 |
134,5 |
250 |
6 |
136 |
270 |
7 |
137,5 |
290 |
8 |
139 |
310 |
9 |
140 |
320 |
10 |
140,5 |
340 |
11 |
142 |
360 |
12 |
142,5 |
390 |
13 |
142 |
395 |
14 |
141 |
420 |
15 |
140 |
450 |
16 |
139 |
490 |
17 |
138 |
520 |
18 |
138 |
560 |
19 |
138,5 |
580 |
Зависимость изменения электропроводности растительной ткани (10 КГц)